
研究紹介
光受容タンパク質の分子生物物理学
1)タンパク質=分子機械の仕組みを知りたい!
 細胞内で何らかの役割を果たしている全てのタンパク質は、形を段階的に変えながら、それぞれの機能を発現しています。その様子は「分子機械」と表現されることがあります。私たちは、タンパク質が、なぜ機械のように働けるのか、その仕組みに興味を持っており、仕組みを明らかにすることを目指した研究を行なっています。こういった仕組みを調べる上で、非常に便利な性質を持っているのが「光受容タンパク質」と呼ばれるタンパク質群です。私たちは、光受容タンパク質の中でも、微生物が持っている「ロドプシン」というタンパク質に、特に魅力を感じていて、主な研究対象としています。このロドプシンは、特に、微生物ロドプシンと呼ばれています。
細胞内で何らかの役割を果たしている全てのタンパク質は、形を段階的に変えながら、それぞれの機能を発現しています。その様子は「分子機械」と表現されることがあります。私たちは、タンパク質が、なぜ機械のように働けるのか、その仕組みに興味を持っており、仕組みを明らかにすることを目指した研究を行なっています。こういった仕組みを調べる上で、非常に便利な性質を持っているのが「光受容タンパク質」と呼ばれるタンパク質群です。私たちは、光受容タンパク質の中でも、微生物が持っている「ロドプシン」というタンパク質に、特に魅力を感じていて、主な研究対象としています。このロドプシンは、特に、微生物ロドプシンと呼ばれています。
2)分子機械としてのタンパク質の魅力
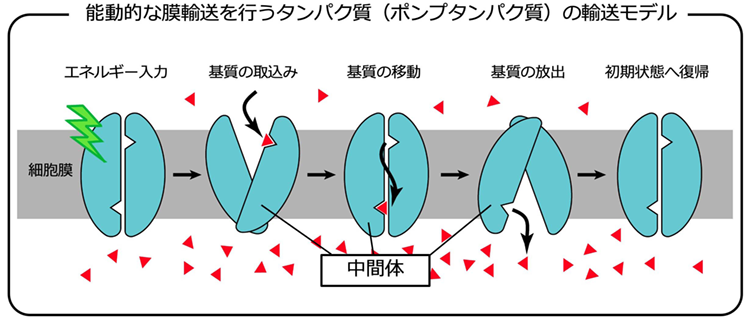
分子機械としてのタンパク質の魅力を、上図に示したタンパク質を例として考えてみます。このタンパク質は、細胞膜を隔てて物質(基質)を輸送するタンパク質(膜輸送タンパク質)です。そのようなタンパク質の中でも、基質を濃度勾配に逆らって輸送できるタンパク質(ポンプタンパク質)を想定しています。微生物ロドプシンの中にも、ポンプとして働くタンパク質があり、私たちの重要な研究対象でもあります。
上の図を見て分かる通り、膜輸送という機能自体は、非常に単純です。基質が、膜の一方から他方へ移動するだけです。しかし、この機能を実現することは容易ではありません。ポンプタンパク質は、ATPの加水分解や光エネルギーのような何らかのエネルギー入力を切っ掛けとして、一連の構造変化を起こします。まず、エネルギー入力を切っ掛けとして、最初にタンパク質の内部を上側の溶液に露出させます。この時、片側の溶液にだけ露出することが重要です。そうでなければ、基質や他の物質のリークを起こしてしまうからです。次に、タンパク質内部に基質を取り込みます。この結合は十分強くなければ、効率よく基質を輸送できません。次の構造変化によって、取り込んだ基質をタンパク質内部で移動させます。先ほどは、強く基質を取り込んだのに、この段階ではその結合を弱めて、タンパク質内部で移動させることができるのです。その次の構造変化を利用して、タンパク質内部を反対側の溶液に露出させ、次いで、そちら側の溶液に基質を放出します。基質を取り込む時とは異なり、こちら側での基質結合は十分弱くないといけません。そうでなければ、やはり効率よく基質を運べないためです。最後にもう一度構造変化を起こして、元の状態に復帰します。基質を一方向(図の上から下へ)輸送するためには、これらの一連の構造変化を、必ず、この順番で行う必要があります。一つでも順番が狂うと輸送できません。タンパク質は、このような規則正しい動きが出来るということです。
タンパク質は、アミノ酸が繋がってできた1本の鎖です。それが折れ畳まれて、機能を持ったタンパク質が出来上がります。そこへ、ひとたび、エネルギーが入力されると、それ以後は、図に描いたような構造変化を、勝手に、かつ、規則正しく行なって、基質を輸送することができます。この反応を何度でも繰り返すことができます。このように、タンパク質は、まさに「分子機械」と呼ぶのにふさわしい性質を持っています。私たちは、勝手に構造が出来上がるタンパク質が、なぜ、機械のように働けるのかに興味を持っています。
3)研究材料として最適な「微生物ロドプシン」
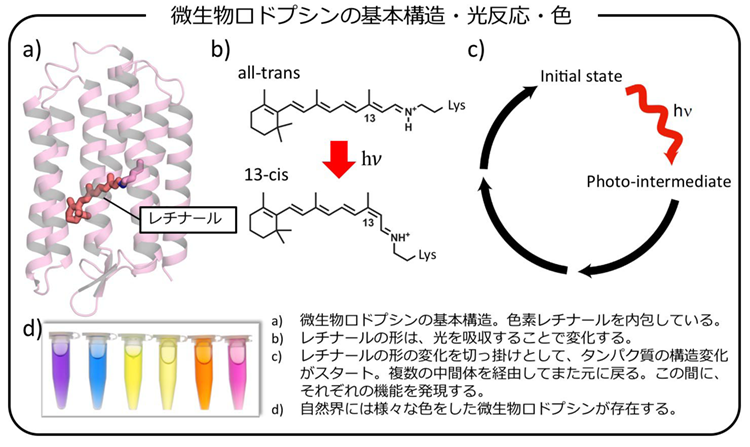
多くの生物は、太陽光をエネルギー源や情報源として巧みに利用しています。この光利用を可能にしているのが「光受容タンパク質」と呼ばれる種類のタンパク質です。全ての光受容タンパク質は、可視光線を吸収するための色素を内包しており、この光吸収を切っ掛けとして、タンパク質側の多段階の構造変化を駆動することができます。その結果、光が来たことを他のタンパク質に伝え、最終的に細胞応答を引き起こしたり、光エネルギーを化学的なエネルギーに変換したりできます。
色素による光の吸収は、一瞬(フェムト秒)で完了します。この光活性化に比べて、後続の構造変化は大変ゆっくり進行します。ですので、時間幅の短い強い光で、光受容タンパク質を一斉に活性化すると、それ以後、これらのタンパク質は、同じタイミングで、同じ状態を経て、反応を進行させていきます。ですので、形が異なる状態(中間体)を、時間を追いながら観測し、それらの中で何が起こっているのかを調べていくことができます。この位相が揃った状態は、光受容タンパク質が光で活性化できるからこそ作れる状態です。他のタンパク質では作れません。そのため、光受容タンパク質は、分子機構の詳細に迫れる非常に優れた研究対象となっています。
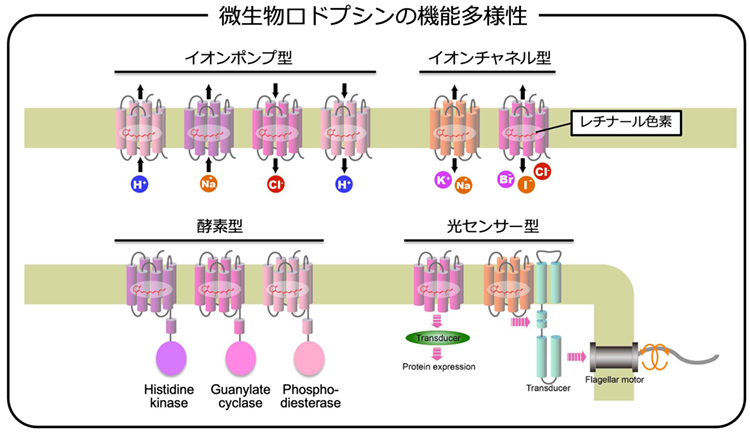
光受容タンパク質の中でも、微生物ロドプシンは、特徴的な地位を確立しています。その名の通り、微生物ロドプシンは、原核生物、真核生物を問わず、様々な微生物に分布している光受容タンパク質です。微生物ロドプシンほど、様々な自然環境に適応した光受容タンパク質はありません。また、この広い分布に対応して、様々な機能を持った微生物ロドプシンが見つかってきています。例えば、a)種々のイオンを能動的に輸送するイオンポンプ型ロドプシン、b)種々のイオンを受動的に輸送するチャネル型ロドプシン、c)光センサーとして働いて、細胞運動を制御するロドプシン、d)光センサーとして働いて、光環境に応じたタンパク質合成を促すロドプシン、e)光でON/OFFする酵素として働いて、光シグナルを増幅するロドプシンなどです。これらの微生物ロドプシンは、全て同じ構造を持っていて、かつ、同じ色素を内包しており、さらに、その色素が起こす構造変化も同じです。それにも拘らず、結果として現れる機能が全く異なります。この性質も、微生物型ロドプシンがもつ魅力の一つです。微生物ロドプシンは、ツッコミどころが満載のタンパク質です。
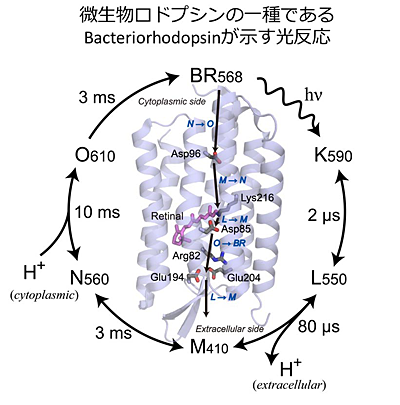
4)私たちの得意技「フラッシュフォトリシス=過渡応答解析」
私たちは、フラッシュフォトリシスという測定方法を得意技としていて、この方法を駆使して、微生物ロドプシンの仕組みを調べています。この方法は、フラッシュ(閃光)によってタンパク質を活性化し、その後に起こるタンパク質側の変化を、時間を追いながら観測する手法です。過渡応答解析と呼ばれる測定手法に含まれます。私たちは、タンパク質の中間体を、それらの色の時間変化(過渡吸収分光測定)や、電気的な性質の時間変化(過渡電気化学測定)で追跡することを得意としています。これらの得意技と、アミノ酸変異導入などの手法を組み合わせて、タンパク質の仕組みに迫る研究を行なっています。

5)最近発表した論文、および、日本語の総説
最近の発表論文とその概要です。一番下の日本語の総説をお読み頂くと、研究の一端をお分かり頂けるかと思います。(2026.05.29更新)
1) アニオンチャネルロドプシン(ACR)のゲート開閉機構を解明:ACRは光によってゲートを開くアニオン(陰イオン)チャネルであり、オプトジェネティクスにおいては、興奮性細胞の活動電位発生を抑制するツールとして使用されています。ゲートの開閉は非常に高速な反応であるため、その詳細な仕組みは謎に包まれていました。本研究では、ゲートが閉まる際にタンパク質内部でプロトン(H+)が移動していることを発見しました。この移動によって生じる電荷の変化が、チャネルの開閉に重要な役割を果たしていることを明らかにしました。
Sudo et al., Proton-Coupled Gate Closing Mechanism in Guillardia theta Anion Channelrhodopsin 1, Journal of Molecular Biology, in press, 169856, 2026
2) H+ポンプロドプシンの環境適応メカニズムを解明:微生物ロドプシンは、自然界のあらゆる環境に生息する微生物が持っています。特に、水素イオンを運ぶ「H+ポンプロドプシン」の適応能力は凄まじく、マイナス30℃以下の極寒の南極海から、70℃を超える灼熱の温泉にまで広がっています。本研究では、異なる温度環境に適応した3種類のH+ポンプについて、光反応を熱力学的に解析しました。その結果、過酷な温度条件下でも効率よく働くための、生命の巧みな生存戦略の一端が見えてきました。
Ohtake et al., Thermodynamic Basis of Temperature Adaptation in Three Outward Proton Pump Rhodopsins Distributed Across Diverse Thermal Environments, Biochemistry, 65, 1314-1324, 2026
3) 世界初、光で動く「有機物トランスポーター」の発見:微生物ロドプシンには、光のエネルギーを使って細胞膜を隔ててイオンを運ぶ「膜輸送」タイプが多く存在します。1970年代に最初のロドプシンが発見されて以来、これまでに輸送が確認されたのはH+やNa+、Cl-といった「無機イオン」だけでした。しかし今回、当研究室は有機イオンを輸送できるロドプシンを世界で初めて発見しました。これは、自然界で見つかった史上初の「光駆動型有機物トランスポーター」であり、これまでの常識を覆す成果です。
Shen et al., Light-Powered Transport of Organic Anions by Microbial Rhodopsins, The Journal of Physical Chemistry Letters, 16, 10528-10535, 2025
4) Cl-ポンプロドプシンの輸送を支える「隠れた重要アミノ酸」を特定:当研究室の先行研究により、塩化物イオン(Cl-)を運ぶ「Cl-ポンプロドプシン」の通り道は特定されていました。しかし不思議なことに、その経路はイオンを運びにくい「疎水的(水になじまない)」なアミノ酸ばかりで構成されていました。そこで、この経路を作るアミノ酸を一つずつ詳しく調べたところ、イオンと直接触れ合うことはないものの、経路の形を適切に維持している2つの重要なアミノ酸を特定しました。これらがストッパー役の別のアミノ酸にCl-が捕まるのを防ぐことで、スムーズな輸送を実現していることが分かりました。
Zhai et al., Important amino acid residues in the chloride pump halorhodopsin that accelerate ion transport despite no direct interaction with the substrate, The Journal of Biological Chemistry, 301, 110703, 2025
5) H+ポンプ型ロドプシンの生理的意義に関する研究:かつて、ロドプシンをもつ微生物は、塩湖に生息する古細菌だけであると考えられていたが、2000年以降、さまざまな環境に生息する多様な微生物が、ロドプシンを持つことが明らかとなった。特に、H+ポンプ型ロドプシンについては、海洋性バクテリアの70%以上が保持するといわれるくらい、普遍的に存在することがわかっているが、細胞における存在意義は完全には証明されてきていない。本研究では、南極に生息する好冷菌を試料として、それが発現するH+ポンプが、光を細胞内エネルギーに変換することや、光照射下で増殖を加速させていることを明らかとした。
Kondo et al., Contribution of Proteorhodopsin to Light-Dependent Biological Responses in Hymenobacter nivis P3T Isolated from Red Snow in Antarctica, Biochemistry, 63, 2257-2265, 2024
インタビュー記事(研究院の広報担当の徂徠さんにインタビューしていただきました)
6) Cl-ポンプ型ロドプシン(通称NpHR)の論文:光励起されたNpHRは、種々の光反応中間体を経由して、元の状態に戻る。この間に、細胞の外側から内側へ能動的にCl-を輸送する。この反応に現れる中間体のうち、どの中間体のときにCl-を放出し、次いで取り込むかを明らかにした論文。当研究室独自の方法であるイオン選択膜を利用した測定系を使って、1回の光反応(約100 msec)の間に、Cl-放出や取り込みに伴って起こる微小なCl-濃度変化を検出した。この結果から、新たなCl-結合サイトがみつかり、さらに、蛋白質内のCl-輸送経路も推定できた。
Hamada et al., Direct detection of the chloride release and uptake reactions of Natronomonas pharaonis halorhodopsin, J. Biol. Chem. 300, 107712, 2024
7) Na+ポンプ型ロドプシン(通称NaR)の論文:光励起されたがNaRが、Na+を放出する際に起こす構造変化の道筋を明らかにした論文。「ピタゴラスイッチ」のような連続的な構造変化が、NaRの中で起こっている!
Sato et al., Multistep conformational changes leading to the gate opening of light-driven sodium pump rhodopsin, J. Biol. Chem. 299, 105393, 2023
インタビュー記事・動画(研究院の広報担当の徂来さんにインタビューしていただきました)(いいね!KOBUNSHI)
8) アニオンチャネルロドプシン(通称ACR)の論文:ACRは、細胞膜に埋まっている膜貫通部分の他に、大きな水溶性ドメイン(通称CPD)を持っている。CPDには、膜貫通部分のアニオン選択性を制御する役割があることを、初めて明らかにした論文。この選択性によって、窒素をNO3-として細胞内に取り込ませる役割があるようだ。
Ohki et al., The preferential transport of NO3− by full-length Guillardia theta anion channelrhodopsin 1 is enhanced by its extended cytoplasmic domain, J. Biol. Chem. 299, 105305, 2023
インタビュー記事・動画(研究院の広報担当の徂来さんにインタビューしていただきました)(いいね!KOBUNSHI)
9) 当研究室が発見したCl-ポンプロドプシン(通称MrHR)に関する論文(当研究室としては3本目の原著論文):野生型のMrHRは、Cl-とBr-しか輸送できないが、イオンの入り口付近のアミノ酸に適切な変異を加えると、SO42-まで輸送できることを示した論文。もっと変異を加えると、もっと大きなイオンを運ばせられるかも!?
Doi et al., Mutations conferring SO42- pumping ability on the cyanobacterial anion pump rhodopsin and the resultant unique features of the mutant, Sci. Rep. 12, 16422, 2022
10) H+ポンプ型ロドプシン(H+ポンプ)に関する論文:H+ポンプは細胞の内側から外側へH+を排出し、細胞が間接的に光エネルギーを利用できるようにしている。H+の輸送経路には疎水的な部分があるが、それにも拘らず、電荷をもったH+を高速で輸送できる。この高速輸送を実現する仕組みには2通りが存在し、それに従って、H+ポンプも2グループに大別できることを示した論文。
Sasaki et al., Replaceability of Schiff base proton donors in light-driven proton pump rhodopsins, J. Biol. Chem. 297, 101013, 2021
11) Na+ポンプ型ロドプシン(通称NaR)に関する論文:NaRに限らず、全ての膜輸送タンパク質は、タンパク質内部を細胞質側溶液と細胞外側溶液に交互に晒して、基質の取り込みと放出を行うはずだが、それらの状態が、リアルタイムで、かつ、区別されて検出されたことはなかった。フラッシュフォトリシスという当研究室の得意な測定方法によって、これらの状態の検出に成功し、それぞれの特徴を明らかにしたという論文。
Kato et al., Real-time identification of two substrate-binding intermediates for the light-driven sodium pump rhodopsin, J. Biol. Chem. 296, 100792, 2021
12) アニオンチャネルロドプシン(通称ACR)の論文:藻類から発見されたACRは、光で活性化されるとアニオンチャネル活性を示すが、生理的な役割は十分には解明されていない。本研究では、藻類の1種 P. sulcataのもつACRが、Cl-やBr-よりも、イオン径の大きいNO3-を強く運ぶことを明らかにした。さらにこのイオン選択性を担っているアミノ酸を特定し、変異導入によって、イオン選択性を消失させることに成功した。ACRは窒素をNO3-として細胞内に取り込む役割があるのかも。
Kikuchi et al., Preference of Proteomonas sulcata anion channelrhodopsin for NO3− revealed using a pH electrode method, Sci. Rep. 11, 7908, 2021
13) 当研究室で発見したCl-ポンプ型ロドプシン(通称MrHR)の興味深い特徴を紹介した日本語の総説:全ての微生物ロドプシンは、H+ポンプ型から機能分化したと考えられている。我々が発見したMrHRはCl-ポンプとして機能するが、H+ポンプにとって重要なアミノ酸も保存しており、たった1アミノ酸を置換しただけで、H+ポンプとして機能し始めた。そのため、MrHRは、H+ポンプから機能分化して間もないCl-ポンプであると考えられる。その後の研究で、さらに成熟したCl-ポンプになるための道筋の一端も明らかにした。
菊川峰志, Cl–ポンプロドプシンの分子機構とH+ポンプへの機能変換, 生物物理 61, 12-15, 2021

光生物学研究室<菊川研究室>
北海道大学 大学院生命科学院
ソフトマター専攻
〒060-0810 札幌市北区北10条西8丁目
理学部2号館 8階 806 / Tel; 011-706-3435
Copyright(c) Hokkaido University Kikukawa Laboratory.
All rights reserved.